森林生态学讲稿第五章森林群落中的种间相互关系.docx
《森林生态学讲稿第五章森林群落中的种间相互关系.docx》由会员分享,可在线阅读,更多相关《森林生态学讲稿第五章森林群落中的种间相互关系.docx(12页珍藏版)》请在冰点文库上搜索。
森林生态学讲稿第五章森林群落中的种间相互关系
群落中的种间相互尖系
一、群落中的种间尖系类型
群落中种与种的尖系总的来说,可分为三种情况,即有利的作用(+)、有害的作用(・)
和没有明显效果的作用(0)。
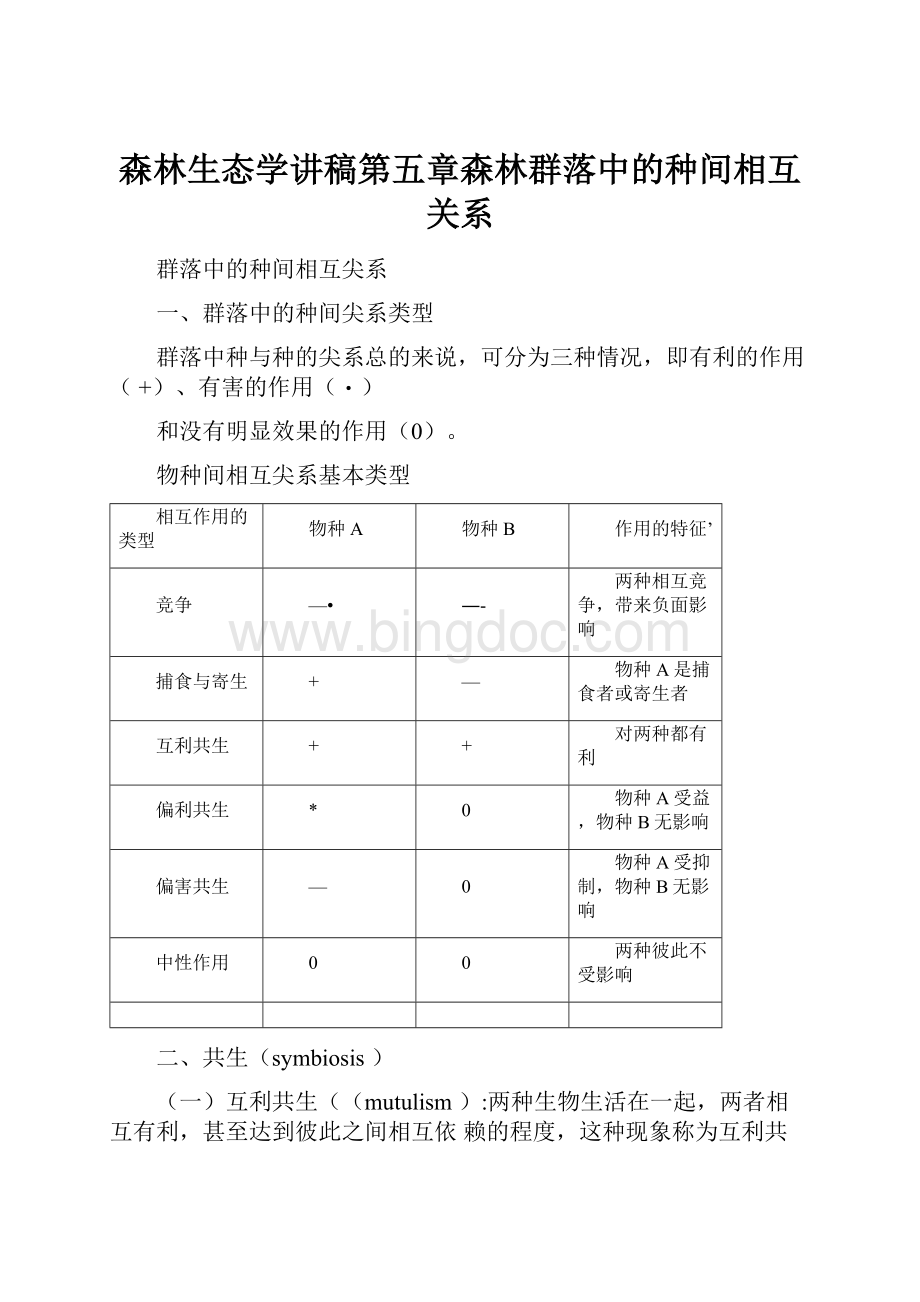
物种间相互尖系基本类型
相互作用的类型
物种A
物种B
作用的特征’
竞争
—•
―-
两种相互竞争,带来负面影响
捕食与寄生
+
—
物种A是捕食者或寄生者
互利共生
+
+
对两种都有利
偏利共生
*
0
物种A受益,物种B无影响
偏害共生
—
0
物种A受抑制,物种B无影响
中性作用
0
0
两种彼此不受影响
二、共生(symbiosis)
(一)互利共生((mutulism):
两种生物生活在一起,两者相互有利,甚至达到彼此之间相互依赖的程度,这种现象称为互利共生。
自然界中生物之间互利共生的现象非常普遍,形式也多种多样,又分为三类:
1连体互利共生
两种生物长期接触,紧密结合在一起的共生尖系。
地衣、菌根、根瘤等都是连体互利共生的典型例子。
一方能吸收水分和无机养分,另一方进行光合作用,提供碳水化合物等有机物。
地衣:
藻类和真菌的共生体,藻类进行光合作用,菌丝吸收水分和无机盐,两者结合、相互补充,共同形成一个统一的整体,生活在岩石或树干这样严酷的环境条件。
根瘤:
固氮菌和豆科植物等根系的共生。
叶瘤也是一种互利共生
菌根是真菌和高等植物根系的共生体,如松树等,在人工造林时(尤其是在养分贫乏的土壤中)接种菌根可显著提高成活率等。
2非连体互利共生(原始合作)
两种生物不是长期结合在一起,而只是间断性接触的共生尖系。
授粉是典型的非连体互利共生尖系的例子。
种子散布是另一类动物与植物之间的非连体共生例子,种子传播者包括食水果动物,它们摄食新鲜水果,但排除或去除种子,热带森林中75%的树种生产新鲜水果,其种子由动物散布。
3防御性互利共生
互利共生的一方能为另一方提供对捕食者或竞争者的防御作用。
蚂蚁一植物互利共生很普遍。
许多植物在树干或叶子上有称做花外蜜腺的特化腺体,为蚂蚁
提供食物源,该腺体分泌富含蛋白质和糖的液体。
在许多种金合欢树中,蚂蚁也通过生活在树的空隙中得到物理保护。
蚂蚁为其宿主提供对抗草食者很强的防御,并且有力地进攻任何入侵者。
(二)偏利共生(commensalism)对一种有利而对另一种无害的共生尖系。
附生,即一种植物定居在另一植物体的表面,附生植物与被附生植物只在定居的空间上发生联系,它们之间没有营养物质的交流。
在热带和亚热带森林中,附生在树木上的地衣、苔鋅、蕨类、兰花、仙人掌等植物,它们本身能自养,通过自身的光合作用制造自己所需的有机养料,所需的矿质元素、水分从降水、尘埃和腐烂树皮中获得。
一些攀缘藤本植物,本身茎不能直立,利用其它的树干作为机械支撑,从而获得更多的光照。
附生可能会转化为其它的尖系。
三、寄生(parasitism)寄生是指一个种(寄生者)寄居于另一个种(寄主)的体内或体表,以寄主的身体为生活空间,并靠吸取寄主的营养而生活o寄生者包括病毒、细菌、真菌、寄生性动物和寄生性种子植物。
寄居于体内的称为体内寄生,寄居于体外的称为体外寄生。
寄生性种子植物分为全寄生和半寄生两类。
全寄生植物从寄主摄取全部营养物质(无叶绿素,不能进行光合作用,其营养全部来源于寄主植物),如菟丝子;而半寄生植物尚能进行光合作用制造有机养分,但只从寄主摄取无机盐类和水分,如桑寄生。
四、异株克生(allelopathy)
德国学者Molich(1937)年提出的,也称他感作用或生物化学作用,通常指一种植物在代谢过程中向体外分泌生物化学物质,对其他植物产生直接或间接的影响。
植物的各类器官,包括根、茎、叶、果、花、种子等可以释放出乙醇、有机酸、醸、醛、酮等。
释放的方式包括淋洗作用、植物残体分解、根系分泌、挥发、树液等。
五、竞争
竞争--是指在同种(种内竞争)或异种(种间竞争)的两个或多个体间,由于它们的需求或多或少地超过了当时空间或共同资源供应状况,从而发生对环境资源和空间争夺的现象。
(一)种内竞争
1林木分化:
在森林生长发育过程中,同一树种年龄相同的植株在形态上和生活能力方面的差异,称为林木分化。
2自然稀疏:
郁闭的林分随着的年龄的增长,单位面积林木株数不断减少的现象。
(1)有自疏和他疏,其中自疏是指同一种群随着年龄的增长和个体的增大,种群的密度
降低的现象。
但是在这种情况下,单位面积的生物量与密度无尖,特定面积能够维持的单位面积产量是恒定的,资源的可利用率决定了最终的生物量,这称为“产量恒定法则”。
他疏是指不同植物种的种群之间,因某一种群密度增加和个体增长,引起另一种群个体死亡的现象,涉及不同物种群之间的竞争。
(2)自疏过程中植物种群密度与个体大小的矢系(作图解释)
1植株的平均大小(重量)随时间而增加;
2只有种群达到自然稀疏线时,才会产生密度制约的死亡,死亡现象在密集种群比在稀疏种群中产生的早;
3具有相同平均重量的林木,在稀疏种群中其年龄比在密集种群中的年轻(由t表
示);
4密集种群和稀疏种群最终达到重量增长和死亡处于平衡的阶段时,总的植物总量
就不再增长。
3最后产量衡值法则和・3/2幕自疏定律(Yoda定律):
最后产量衡值法则:
在一定范围内,当条件相同时,不管初始播种密度如何,最后产量差不多都是一样的5即最后产量衡值法则(lawofconstantfinalyield)
最后产量衡值法则可用下式表示:
”二Wxd=7C-
丫,单位面积产量;w植物个体平均重量;d,密度;Ki,常数
・3/2幕自疏定律(经验公式,非理论推导):
日本学者Yoda发现植物的平均干重(w)和存活个体密度(d)之间的相互尖系可表示为:
两边取对数:
=lgC-a翻
式中C和a是常数,a是密度和植物平均干重的对数曲线的斜率logC是曲线在纵坐标上的截距。
不同植物的C值约在3.5-4.5之间,而a=3/2。
因此W=Cd-3/2被称为・3/2自疏法则。
■3/2幕自疏定律的前提(假设条件)在密集而生长旺盛的同龄植物种群中,如果在
没有其他物种的竞争和非密度制约因子(如火、干旱、疾病等)的胁迫下,个体的死亡或稀疏是由种内竞争引起的,在稀疏过程中平均个体质量与种群密度间存在着幕指数为・3/2的幕函数尖系。
此尖系式还适用于林分中的林木平均单株树干材积(V)与最大密度(dm)之间的尖系:
日本几个树种平均单株干材积(V)的最大密度(dm)线如下:
桜卩杉K527x114
扁柏J-517X
落口i松V1.046X5。
(二)种间竞争
1类型
(1)资源利用性竞争(exploitationcompetition):
两种生物间因资源总量减少而产生的对竞争对手生长、存活和繁殖的间接影响,即两种生物同时竞争利用同一种资源。
(2)相互干涉性竞争(interfereneecompetition)指竞争个体间直接的相互作用,即一种
生物借助行为排斥另一种生物使其得不到资源。
最明显的例子是动物为了竞争领域或食物而进行的打斗。
树冠磨擦也是一种。
2特征
(1)不对称性,即竞争各方影响的大小和后果不同。
即竞争后果不等性。
(2)共轨性,即对一种资源的竞争,能影响对另一种资源的竞争结果。
3竞争制E斥原理(competitiveexclusion)
又称为高斯(前苏联生态学家)假说(Gausehypothesis,即在一个稳定的环境中,两个受资源限制的但具相同资源利用方式的种,不能经久的共存一处。
换言之完全的竞争者不能共存。
高斯(Gause)以两种分类上和生态上很接近的草履虫为实验材料,直接观察两个种的竞争结果。
他取双小核草履虫和大草履虫两个种相等数目的个体,用一种杆菌作为饲料。
当单独培养时,两种均呈“S增长曲线,但当两种混合培养时,即同时放在一个基本恒定的环境里培养时,开始两种都增长,随后双小核草履虫数量增加,而大草履虫个体数下降,并于16天后,只有双小核草履虫生存,而大草履虫完全灭亡
4洛特卡■沃尔泰勒(Lotaka-Volterra)种间竞争模型
(1)竞争模型:
该模型是在Logistic方程的基础上建立起来的,假定有两个物种,当它们单独生长时其增长形式符合逻辑斯谛模型,其增长方程是:
阿K.・Ni
种群1
卡=汎(晋种群2
式中N1、N2——分别为两个物种的种群数量;
心、K2-分别为两个物种种群的环境容纳量;
n、r2——分别为两个物种种群增长率。
如果将这两种放置在一起,则它们就要发生竞争,从而影响种群的增长。
兽-曲巴种群1竞争方程
警匚沁(*)种群2竞争方程
2
式中a表示物种1的环境中,物种2对物种1竞争系数;B表示物种2的环境中,物种1对物种2的竞争系数。
(2)竞争系数a意义分析
1表示物种1的环境中,每存在一个物种2的个体,对于物种1种群的效应,即一具物种2个体所利用的资源相当于a个物种1的个体;
2若0=1,表示每个物种2个体对物种1种群所产生的竞争抑制效应,与每个物种1个体对自身种群所产生的效应相等;
3若a>1,表示每个物种2个体对物种1种群所产生的竞争抑制效应,大于物种1个体对自身种群所产生的效应;
4若a<1,表示每个物种2个体对物种1种群所产生的竞争抑制效应,小于物种1个体对自身种群所产生的效应
(3)竞争结局结果
从理论上讲,两个种群在一起竞争时,可能产生以下的结局
勺邙岭Ki0皿
⑶
0
K]
"dT
Sb
两亍种群之间竞争可能产生的四种结局
⑴坷取胜⑵叫取胜⑶稳定的平衡俩种共存)(4)不稳定的平衡(两种都有可能取胜)(倩SmitklPSO)
1种群1取胜,种群2被挤掉。
该情况发生在Ki>K2/B,K2由于©,K2/B右边这个面积内,种群2已超过最大容纳量而不能增长,而种群1仍能继续增长,因此,种群1取胜。
2种群2取胜,种群1被挤掉。
其情况与
(1)相反,该情况发生在K2>Ki/0而KkK2/B的时候。
在匕、Ki/0Ki、K2/B这块面积内,种群1不能增长,而种群2能继续增长。
因此,种群2取胜。
3表示两个种群共存,形成稳定的平衡局面。
该情况产生在KkK2/B和K2/a的时候,两条对角线相交;其交点E即为平衡点。
由于KkK2/B在三角形K1EK2/B中,种群1不能增长,而种群2能继续增长,箭头向平衡点收敛。
同样,因为K2<心/a,在三角形Ki/aEK中,种群2不能增长,而种群1增长,箭头也向平衡点收敛,从而形成稳定的平衡。
4当Ki>K2/BK2>K〃a时,两条对角线相交,出现平衡点,但它是不稳定的。
因为Ki>K2/B在三角形K1EX2/B中,种群2不能增长,种群1能增长,箭头不收敛。
同样因为K2>Ki/須在三角形K2E*K/a中,种群1不能增长,种群2能增长,箭头也不能收敛。
因此,平衡是不稳定的。
(4)竞争结局分析
1/Ki和1/K2这两个值,可以被视为种群1和种群2的种内竞争强度指标。
其理由在于在一个空间中,如能“装下”更多的同种个体,则其种内竞争就相对的小。
因此,1/K值的大小可以作为种内竞争强度的指标。
同理,/©值可以被视为种群2对种群1的种间强度指示,而a/Ki则为种群1对种群2的种间竞争强度指标。
因此,竞争的结局取决于种内竞争和种间竞争相对大小。
如种群的种间竞争强度大,而种内竞争强度小,该种群取胜。
反之,若某种群的种间竞争强度小,而种内竞争强度大,则该种群竞争失败。
若两个物种的种内竞争均比种间竞争强烈,两物种就可能会稳定共存;若种间竞争均比种内竞争强烈,那就不可能有稳定的共存。
5生态位(niche)理论
(1)生态位的概念及研究史生态位主要指在自然生态系统中一个种群在时间、空间上的位置及其相矢种群之间的功能尖系(怀梯克,1975)o
空间生态位:
生态位一词最早由格林尼尔(Grinell)于1917年提出,用来表示划分生境(habitat)的空间单位和一个物种在生境中的地位。
他认为生态位是一个物种所占有的生境。
实际上,强调的是空间生态位(spatialniche)的概念。
营养生态位:
英国生态学家埃尔顿(Elton)(1927)将生态位看作是“物种在生物群落中的地位与功能作用”他强调是物种之间的营养尖系,实际上指的是营养生态位
(trophicniche)的概念。
超体积生态位:
美国生态学家哈钦森(Hutchinson”1957)从空间、资源利用等方面考虑提出了n维生态位(n-dimensionalniche)的概念发展了生态位概念。
因为环境变量是多维的(三维变量成为体积)5所以把n维生态位称为超体积生态位(hyper・volumeniche),假设影响有机体的每个条件和有机体能够利用的每个资源都可被当作一个轴或维,在此轴或维上,可以定义有机体将出现的一个范围,同时考虑一系列这样的维,就可以得到有机体生态位的一个增强了的定义图。
基础生态位和现实生态位:
哈钦森还认为,生态位应该区分为两种:
基础生态位和现实生态位。
基础生态位(fundamentalniche)代表不存在其他种竞争的情况下该种能占据的最大生态位。
而现实生态位(realizedniche)仅是基础生态位的一部分,是指该种在存在有其他种竞争的情况下所占据的实际部分,竞争种类越多,使某物种占有的实际生态位可能越来越小。
美国学者环梯克(Whittaket)(1975)认为,生态位是每个种在一定生境的群落中都有不同于其他种的自己的时间、空间位置,也包括在生物群落中的功能地位,并指出生态位的概念与生境和分布区的概念是不同的,生境是指在生物生存的周围环境,分布区是指种分布的地理范围,生态位则说明在一个生物群落中某个种群的功能地位。
生态位越接近,重叠越多,种间竞争就越激烈,将导致一物种灭亡或生态位分离。
总之,种内竞争促使两物种生态位接近,种间竞争又促使两物种生态位分离,这是两个相反的
进化方向0
6用竞争排斥原理与生态位理论解释自然生物群落:
(1)一个稳定的群落中占据了相同生态位的两个物种,其中一个终究要灭亡;
(2)一个稳定的群落中,由于各种群在群落中具有各自的生态位,种群间能避免直接的竞争,从而保证了群落的稳定;
(3)群落是一个相互起作用的、生态位分化的种群系统,各种群在它们对群落的时间、空间和资源的利用方面,以及相互作用的可能类型,都趋向于相互补充而不是直接竞争。
大家配合共同生活,更有效地利用环境资源,从而保证了群落在一个较长时间有较高的生产力,具有更大的稳定性。
(4)竞争可以导致多样性而不是灭绝,竞争在塑造生物群落的物种构成中发挥着主要作用。
竞争排斥在自然开放系统中,很可能是例外而不是规律,因为,物种常常能够转换它们的功能生态位去避免竞争的有害效应。
(第4点可不要)
六、捕食(Predation)作用
1.概念:
即一种生物摄取其他种生物个体的全部或部分为食的矢系。
狭义的捕食是指食肉动物吃食草动物或其它食肉动物。
广义的捕食也包括植食(或草食)。
2.捕食的生态意义
(1)捕食可调节猎物种群的数量,限制其分布。
(2)捕食可改善猎物种群的质量,是一种主要的选择压力;
(3)捕食现象是影响群落结构的主要生态过程。
3协同进化(coevolution)
在种间相互作用的影响下,不同生物间相尖性状在进化中得以形成和加强的过程。
捕食者对策:
动物为了获得最大的觅食效率会采用种种方法和措施:
(1)在形态学上,如捕食者在进化过程中发展了锐齿、利爪、尖喙、毒牙等工具;
(2)在行为上,运用诱饵、追击'集体围猎等方式,以便有利地捕食猎物;
反捕食策略:
(1)隐藏:
动物利用保护色、警戒色、拟态、假死或地形、草丛和隐藏所等;
(2)逃避:
逃回洞穴、逃回树上或跳入水中、奔跑等;(3)自卫:
在动物发现敌害后无处隐藏,又来不及逃避时所采取的一种反捕食策略。
集体抵御等。
植物看上去是只有坐以待毙之能,但是实际上,植物也有自己丰富多采切行之有效
的防御手段。
植物以两种主要方式来保护自己免遭捕食:
①毒性与差的味道;②防御结构,在植物王国已发现大量的多种化学武器来保卫植物免遭捕食者和寄生者的进攻。
这些次生化合物或直接有毒,或可降低植物的食物价值,如降低动物肠道对植物叶组织蛋白的吸收。
防御结构在各种水平上都存在,从叶表面可陷住昆虫及其他无脊椎动物的微小绒毛,到可阻止哺乳类食草动物的大型针刺。
经历过落叶的植物,其次生化合物水平及防御结构大小都会提高或“被诱导”。
 升级会员
升级会员 