克隆载体与表达载体.docx
《克隆载体与表达载体.docx》由会员分享,可在线阅读,更多相关《克隆载体与表达载体.docx(26页珍藏版)》请在冰点文库上搜索。
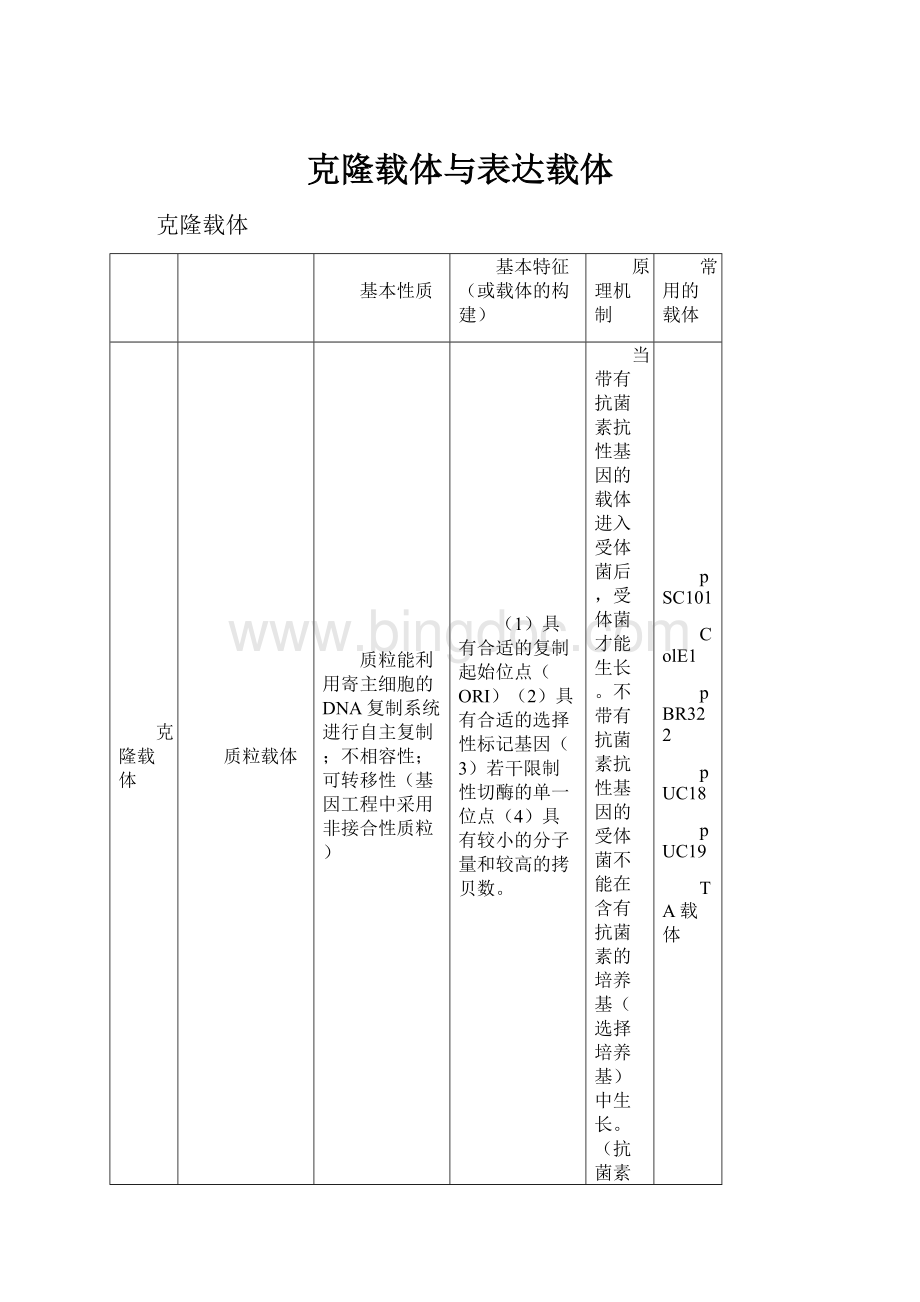
克隆载体与表达载体
克隆载体
基本性质
基本特征(或载体的构建)
原理机制
常用的载体
克隆载体
质粒载体
质粒能利用寄主细胞的DNA复制系统进行自主复制;不相容性;可转移性(基因工程中采用非接合性质粒)
(1)具有合适的复制起始位点(ORI)
(2)具有合适的选择性标记基因(3)若干限制性切酶的单一位点(4)具有较小的分子量和较高的拷贝数。
当带有抗菌素抗性基因的载体进入受体菌后,受体菌才能生长。
不带有抗菌素抗性基因的受体菌不能在含有抗菌素的培养基(选择培养基)中生长。
(抗菌素选择原理)
pSC101
ColE1
pBR322
pUC18
pUC19
TA载体
噬菌体载体
λ噬菌体
其DNA两端的5′末端各带有12个碱基的互补单链,即cos位点。
有可替代区,即非必需区。
用外源DNA片段替代这个区域,不会影响噬菌体颗粒的形成。
(1)基因组大小;去除非必需区,建立外源DNA片段的克隆或替换位点
(2)在DNA的非必需区插入选择标记:
lacZ基因;基因cⅠ失活(cI基因:
溶源过程控制基因);Spi筛选(野生型λ噬菌体在带有P2原噬菌体的溶源性E.coli中的生长会受到限制的表型,称作Spi+,即对P2噬菌体的干扰敏感)
1)通过裂解过程增殖载体2)载体与外源DNA的酶切3)外源DNA与载体的连接4)重组噬菌体的体外包装5)包装噬菌体颗粒的感染6)筛选(λ噬菌体载体的克隆原理)
插入式载体
置换型载体(取代型载体)
M
13噬菌体载体
M13噬菌体的基因组为单链DNA。
噬菌体颗粒的大小受其DNA端点制约的,不存在包装限制。
只感染雄性大肠杆菌
基因间隔区(intergenicregion,IG区)基因II与基因IV之间存在一段507bp的基因间隔区,含有复制起始位点,是实施改造、构建人工载体的重点区域。
②IG区只有一个BsuI切点。
(2)加入酶切位点,在IG区加入单一切酶位点。
M13mp1在IG区插入一个大肠杆菌的LacZ’(-肽序列)。
使克隆的DNA片段以特定单链的形式输出受体细胞外,M13重组分子筛选简便,被M13噬菌体感染的受体细胞生长缓慢,形成混浊斑,易于辨认挑选。
而且重组分子越大,混浊斑的混浊度亦越大但M13-DNA载体的最大缺陷是装载量小,只有1.5kb
1、以(+)链DNA为摸板,合成互补(-)链,该双链称复制型DNA(RFDNA)。
2、RFDNA在宿主细胞能快速增殖,可增加到每个细胞约200个拷贝。
3、单链特异的DNA结合蛋白结合在(+)链上,从而阻断了其互补链,即(-)链的合成,这样,细胞就会不断的合成(+)链DNA4、游离出来的(+)链DNA先与基因V的编码产物形成特异的DNA-蛋白质复合物,然后转移到寄主细胞膜,同时基因V的蛋白质从(+)DNA链上脱落下来,余下的M13(+)链DNA则是从其感染的寄主细胞的细胞膜溢出的过程中,被外壳蛋白包装成病毒颗粒的。
(M13噬菌体产生单双链DNA的机制)
M13mpnn代表系列数字②对M13mp1的改进加上常用的酶切位点。
M13mp2:
LacZ’5’端的第13个核苷酸G突变成A,产生了一个EcoRI切点
噬菌体-质粒杂合载体
黏粒载体
考斯质粒是一类人工构建的含有λ-DNAcos序列和质粒复制子的的特殊类型载体。
能像-DNA那样进行体外包装,并高效转染受体细胞;能像质粒那样在受体细胞中自主复制具有较高容量的克隆能力:
45kb;具有与同源性序列的质粒进行重组的能力
粘粒(cosmid)是带有cos序列的质粒。
cos序列是噬菌体DNA中将DNA包装到噬菌体颗粒中所需的DNA序列。
黏粒的组成包括质粒复制起点(colE1),抗性标记(ampr),cos位点,因而能象质粒一样转化和增殖。
克隆的最大DNA片段可达45kb。
有的粘粒载体含有两个cos位点,在某种程度上可提高使用效率。
设计构建的柯斯质粒一般长4~6kb。
其上的cos位点的一个重要作用是识别噬菌体的外壳蛋白。
凡具有cos位点的任何DNA分子只要在长度相当于噬菌体基因组,就可以同外壳蛋白结合而被包装成类似噬菌体λ的颗粒。
因此,插入柯斯质粒的外源DNA可大于40kb。
重组的柯斯质粒可象噬菌体λ一样感染大肠杆菌,并在细菌细胞中复制。
噬菌粒载体
能像质粒那样在受体细胞中自主复制能像M13-DNA那样体外包装,并高效转染受体细胞装载量比常规的M13mp系列要大很多(10kb通过克隆双链DNA能获得同等长度的单一单链DNA
重组操作简便,筛选容易
噬菌粒载体综合了质粒载体和M13噬菌体载体优点,含有ColEl复制起始位点、抗生素抗性选择标记和丝状体噬菌体DNA间隔区(含有噬菌体DNA复制起始、终止以及噬菌体颗粒形态发生所必需的全部顺式作用元件)
它具有质粒的复制起点、选择性标记、多克隆位点等,方便DNA的操作,可在细胞稳定存在;又具有单链噬菌体的复制起点,在辅助噬菌体的存在下,可进行噬菌体的繁殖,产生单链的子代噬菌体
1.具有较小分子量基因组DNA,可克隆10kb的外源DNA片段,并易于进行分离与操作;2.编码一个ampr基因作为选择记号,便于转化子的选择;3.拷贝数含量高4.存在着一个多克隆位点区,因此多种不同类型的外源DNA片段,不经修饰便可直接插入到载体分子上;5.多克隆位点区阻断了大肠杆菌lacZ基因的5’-端编码区,可按照IPTG显色反应试验筛选6.lacZ基因是置于lac启动子的控制之下,插入的外源基因便会以融合蛋白质的形式表达;7含有质粒的复制起点,可复制形成大量的双链DNA分子8.带有一个M13噬菌体的复制起点,在有辅助噬菌体感染的寄主细胞中,可以合成出单DNA拷贝,并包装成噬菌体颗分泌到培养基中;9.;在pUC118和pUC119这两个载体中,多克隆位点区的核苷酸序列取向是彼此相反的,于是它们当中的一个可转录克隆基因的正链DNA,另一个则可转录出负链DNA
pUC118和pUC119
人工染色体
染色体具有复制功能,利用染色体的复制元件来驱动外源DNA片段复制的载体称为人工染色体载体
其装载外源DNA的容量比质粒、噬菌体和噬菌体-质粒杂合载体等有很大的拓展,甚至可以跟染色体的大小相媲美。
人工染色体载体拷贝数少,制备困难,通常采取“穿梭载体”的策略来解决
含有质粒载体所必备的第一受体(大肠杆菌)质粒复制起始位点,这样的载体在大肠杆菌可以按质粒复制形式进行高拷贝复制,含有第二受体(如酵母)端粒(TEL)、DNA复制起始位点(ARS)和着丝粒(CEN)以及合适的选择标记。
载体在体外与目的DNA重组后转化到第二受体细胞,按照染色体复制的形式进行复制和传递。
筛选第一受体的克隆子一般采用抗生素抗性选择标记;筛选第二受体的克隆子常用与受体互补的营养缺陷型。
酵母人工染色体载体YAC
细菌人工染色体载体BAC
P1噬菌体人工染色体载体PAC
质粒载体总结
质粒载体
类型
长度
选择标记
克隆位点
pSC101
天然质粒,属严紧型、低拷贝型
9.09kb
四环素抗性Tetr
7个克隆位点:
EcoRI、XhoI、PvuI、HindIII、BamHI、SalI、SamI
主要使用EcoRI
ColE1
天然质粒,属松弛型多拷贝型
6.3kb
大肠杆菌素(colicin)E1和对E1免疫的基因(immE1)
EcoRI位于E1部,插入外源DNA会导致E1失活,使受体菌不能合成E1(ColE1-),但仍然表现出对E1免疫型(ImmE1+)
pBR322
元件来源①复制起点orip
MB1系列(来源于ColE1)的高拷贝型复制起点②Ampr基因pSP2124质粒的Ampr基因③Tetr基因pSC101的Tetr基因。
4363bp
氨苄青霉素和四环素抗性
24个克隆位点。
其中9个会导致Tetr基因失活(如BamHI、HindⅢ、SalI);
3个会导致Ampr基因失活(ScaI、PvuI、PstI)。
pUC系列
(i)来自pBR322质粒的复制起点(ori);(ii)氨苄青霉素抗性基因(ampr),但它不再含有原来的核酸切限制酶的单识别位点;(iii)大肠杆菌β-半乳糖酶基因(lacZ)的启动子及其编码α-肽链的DNA序列,此结构特称为lacZ′基因;(iv)位于lacZ′基因中的靠近5′-端的一段多克隆位点(MCS)区段,但它并不破坏该基因的功能
2.7kb
Ampicillin抗性和lacZ的α肽互补(蓝白斑)相结合。
10个连续的单一限制酶切位点,位于lacZ’基因的5’端。
TA载体
耐热聚合酶可在PCR产物的3’端加上一个非配对的脱氧腺嘌呤核苷(A)。
根据这一特点研制出线性TA质粒载体,其5’端各带有一个不配对的脱氧胸腺嘧啶(T),用该载体可进行PCR产物的直接克隆。
TA载体构建:
在一般质粒载体的基础上构建。
方法1:
先采用限制性切核酸酶酶切使质粒载体线性化,再通过K1enow片段将酶切的线性载体末端补平
方法2:
利用产生平末端的限制性切核酸酶酶切产生平末端,最后将线性化钝末端质粒载体加T反应形成。
目前有很多公司推出了TA质粒载体。
λ噬菌体载体
λ噬菌体
载体
分类
插入式载体
一种只具有一个可供外源DNA插入的克隆位点的派生载体
λ噬菌体载体相对于质粒载体来说,克隆片段较大,所以一般用于cDNA文库或基因组文库的构建。
cI基因插入失活:
如λgt10、λNM1149等载体,在cI基因上有EcoRI及HindIII的酶切位点。
外源基因插入后将导致cI基因的失活。
cI基因失活后将导致噬菌体不能溶原化,产生清晰的噬菌斑。
相反,产生混浊的噬菌斑。
LacZ基因插入失活:
如charon16A载体,在非必须区段引入lacZ基因,在lacZ基因上有EcoRI位点,插入失活后利用X-gal法筛选(蓝白筛选)
置换型载体(取代型载体)
具有成对的克隆位点,在这两个位点之间的λDNA区段可以被外源DNA片段所取代。
野生型噬菌体染色体的中段对于噬菌体的感染和复制是非必要的,外源DNA可以取代这一片段
这些载体是用Lac5替换入噬菌体的中间区段,同时将Lac5作为选择标记
使用时用EcoRI水解,去掉中间的片段,再与欲克隆片段在体外进行重组、包装。
而后,感染E.coli使之在E.coli繁殖,并裂解E.coli,形成空斑。
置换型λ噬菌体是使用最广泛的载体。
人工染色体
类别
元件
优点
酵母人工染色体载体
YAC
是最常用来克隆很大DNA片段(2Mb)的系统。
为构建YAC需要结合四个短序列:
即二个端粒、一个着丝粒和一个ARS元件,并将一适当大小的外源DNA连接成一线性DNA分子,而端粒序列位于正确的末端。
YAC的构建有比在细菌中克隆的一些优点,因为某些真核细胞的序列,特别是重复序列是难以甚至不可能在细菌中繁殖的,酵母细胞却具有这样的能力
细菌人工染色体载体
BAC
是基于大肠杆菌的F质粒构建的,高通量低拷贝的质粒载体
每个环状DNA分子中携带一个抗生素抗性标记,一个来源于大肠杆菌F因子(致育因子)的严谨型控制的复制子oriS(Shizuyaetal.1992),一个易于DNA复制的由ATP驱动的解旋酶((RepE)以及三个确保低拷贝质粒精确分配至子代细胞的基因座(parA,parB,和parC)
①以大肠杆菌为寄主,转化率高,构建BAC文库比YAC文库更容易;②BAC载体以环型超螺旋状态存在,从大肠杆菌中提取质粒较方便,而从酵母中分离DNA较困难;
③BAC的复制子来源于F因子,可稳定遗传,嵌合及重组现象少;④可以通过菌落原位杂交来筛选目的基因,方便快捷;⑤BAC载体在克隆位点的两侧具有T7和Sp6聚合酶启动子,可以用于转录获得RNA探针或直接用于插入片段的末端测序。
表达载体
分类
结构组成及表达元件
特点或优点
备注
质粒表达载体
原核表达载体
1启动子、
2转录终止子(防止克隆的外源基因表达干扰载体的稳定性)
3核糖体结合位点
表达方式有:
组成型表达,
诱导型表达。
融合型表达,
分泌型表达。
启动子类型:
根据作用方式及功能分类①组成型启动子
②组织特异启动子又称器官特异性启动子。
③诱导型启动子
酵母表达载体
来自pBR322基本骨架含有该质粒复制起始位点(ori),lacZ基因、bla或ampr、tetr等基因。
含有URA3、HIS3、LEU2、TRPl、LYS2与特定的宿主营养缺陷突变体互补筛选或利用zeocin、basticidin(杀稻瘟菌素)的抗性基因筛选。
启动子有GAP、AOXl、AUGl和GAL1等。
GAP是组成型启动子。
其余是诱导型启动子。
酵母表达载体多为穿梭载体:
既可以在大肠杆菌中繁殖,获得足够的载体DNA进行体外操作,又可以在酵母中复制、表达和选择。
融合蛋白表达载体:
于外源基因插入位点的C端或N端插入了1个6×His标签,或在5’端插入了α-因子作为分泌信号
由于原核生物和真核生物存在糖基化、酰基化等翻译后修饰反应机制的差异,真核基因的原核表达难以获得有活性的蛋白。
酵母成为真核表达的首选系统。
Ti质粒表达载体
一元载体
一元载体就是含目的基因的中间表达载体与改造后的受体(Ti质粒)通过同源重组所产生的一种复合型载体,也称为共整合载体。
由于该载体的T-DNA区与Vir区紧密连锁,也称为顺式载体(cis-vector)
Ti质粒是根癌农杆菌中发现的可引发植物产生冠瘿瘤的质粒。
根据冠瘿瘤合成的冠瘿碱种类,Ti质粒可分为章鱼碱、农杆碱、农杆菌素和琥珀碱4种不同类型Ti质粒可分为T-DNA、Vir、Con和Ori4个功能区。
T-DNA区是Ti质粒中能转移到植物细胞的区域。
Vir区位于T-DNA区的上游,其表达产物可激活T-DNA向植物细胞的转移,这一个区域也称为致病区。
Con区该区含有与农杆菌之间接合转移有关的基因(tra),这些基因受宿主细胞合成的冠瘿碱激活,使Ti质粒在细菌之间转移。
Ori区调控Ti质粒的自我复制,为复制起始区
构建的基本原理:
引入分子质量较小的中间载体,将欲转化的目的基因及抗性标记基因构建到中间载体上。
将Ti质粒上T-DNA的致瘤基因全部去掉,仅保留其两边界及与T-DNA转移所必需的25个碱基序列,构建成所谓卸甲载体。
在卸甲载体的T-DNA区被删除致瘤基因位点插入中间载体同源的质粒序列,将中间载体转入含有卸甲载体的根癌农杆菌中,构建在中间载体上目的基因可通过同源重组整合到卸甲载体Ti质粒的T-DNA区,并与卸甲载体Ti质粒一起复制。
二元载体
双元表达载体系统主要包括两个部分:
一部分为卸甲Ti质粒,这类Ti质粒由于缺失了T-DNA区域,完全丧失了致瘤作用,主要是提供Vir基因功能,激活处于反式位置上的T-DNA的转移。
另一部份是微型Ti质粒,它在T-DNA左右边界序列之间提供植株选择标记如NPTII基因以及LacZ基因等。
双元载体系统的构建的原理是Ti质粒上的Vir基因可以反式激活T-DNA的转移。
与共整合载体所不同的是,它不依赖两个质粒之间的同源序列,不需要共整合过程就能在农杆菌独立复制
双元表达载体构件方便,转化效率高于一元载体。
病毒表达载体
杆状病毒表达载体
杆状病毒表达系统的基本元件:
(1)启动子:
多个启动子同时表达多个外源基因。
(2)polyA加尾信号:
(3)同源重组序列。
(4)穿梭载体必需元件。
除带有病毒自身的复制起始位点外,还带有pUC质粒的复制子及以ampr,可以在细菌进行扩增。
(5)筛选标记。
①重组蛋白具有完整的生物学功能:
接近天然蛋白。
②能进行翻译后的加工修饰:
研究糖基化对蛋白质结构与功能影响方面的理想模型。
③表达水平高:
最高可使目的蛋白的量达到细胞总蛋白的50%。
④能容纳大分子的插入片段:
上限未知。
⑤能同时表达多个基因:
在同一细胞同时表达多个基因。
⑥能表达基因组DNA:
昆虫杆状病毒表达系统具有剪切的功能。
⑦对重组蛋白进行定位的功能:
如将核蛋白转送到细胞核上,膜蛋白则定位在膜上,分泌蛋白则可分泌到细胞外等。
杆状病毒科病毒,杆状病毒颗粒,双链闭合环状DNA病毒,专一性感染无脊椎动物
将外源基因先克隆到转移载体上,再与病毒基因组共同转染细胞,通过细胞的同源重组获得带有外源基因的重组病毒的策略。
腺病毒载体
腺病毒DNA末端结构突出特点1)带有100bp反向的末端重复序列(ITR),而且在每一条单链的5’-末端还共价地连接着末端结合蛋白,对病毒的复制有重要作用。
2)当它从病毒颗粒中分离出来之时,便会自发地环化起来,尔后许多环形分子又聚合成多聚体
腺病毒载体是目前最为广泛应用的基因载体,也是唯一基因药物的载体双链DNA的分子大小约为36kb.
可应用于1.基因治疗2.表达真核基因3.研制疫苗
首先构建一个含多克隆位点和筛选标志的转移质粒,该质粒含有病毒基因组某段早期序列;然后将一个含有启动子一外源基因-polyA的表达盒插入到上述质粒中腺病毒E1、E3或E4至右侧的ITR区之间,构建成载有外源基因的穿梭质粒。
反转录病毒载体
泡沫病毒类
从5’端开始依次是:
①5’长末端重复序列(5’-LTR),带有增强子和启动子序列;②psi序列,又称ψ序列,是病毒包装所必须的信号序列;③gag,编码病毒衣壳蛋白;④pol,编码反转录酶和整合酶(Int);⑤env,编码病毒颗粒外层包装蛋白;⑥3’长末端重复序列(3’-LTR),带有polyA加尾信号序列。
一类含单链RNA的动物病毒。
它的基因组含有2条相同的正链RNA分子,包装成二倍体病毒颗粒。
(1)在大多数情况下,反转录病毒的肿瘤基因(onc)都能够在正常的细胞中转录。
2)反转录病毒的寄主围相当广,包括无脊椎动物和脊椎动物。
3)反转录病毒具有强启动子,外源基因可得到有效表达。
4)反转录病毒不但感染效率高,而且不招致寄主细胞的死亡
载体的构建:
分离原病毒DNA→删去部分序列→组入选择性标记基因、目的基因和调控元件→克隆到含有大肠杆菌复制起始位点的克隆载体→转化大肠杆菌→获得反转录病毒克隆载体→辅助细胞系(包装细胞系)→扩增
慢病毒类
致癌病毒类
SV40病毒型转化载体
根据SV40DNA进入敏感动物细胞的转方向,分为早期转录区和晚期转录区。
早期转录区包括与病毒感染相关的t抗原基因(smallT-antigen)和T抗原基因(largeT-antigen)等;含有BamHI等限制性切核酸酶识别位点;晚期转录区包括与病毒壳蛋白合成相关的基因,含有EcoRi等限制性切核
酸酶识别位点。
①含有能够被真核细胞识别的有效的启动子。
②有许多种动物病毒,在其感染周期中都能够持续地复制,使其基因组拷贝数达到相当高的水平。
③有些动物病毒具有控制自己复制的顺式元件和反式作用因子。
④有些动物病毒,在它们的复制过程中能高效稳定地整合到寄主核基因组上。
⑤病毒的外壳蛋白质能够识别细胞接受器。
用病毒外壳蛋白质包装重组质粒DNA形成的假病毒颗粒,即构成了一种高效的转化体系
猿猴空泡病毒40(Simianvacuolatingvirus40,SV40)基因组是一种环形双链的DNA,其大小仅有5243bp,很适于基因操作。
导致人体癌变的可能性极低,对人体是安全的。
一些质粒型表达载体带有来自SV40DNA的个别调控区,位于病毒的早期与晚期转录单位之间,包括1、DNA复制起始位点2、早期与晚期mRNA转录的起始位点3、能激活复制起始位点并可自动调节早期转录的T抗原结合位点4、可被细胞转录因子识别的一般由3个G/C丰富的21bp同向复制区组成的序列5、组成SV40早期增强子的2段72bp同向重复序列。
大片段表达载体
TAC-可转移人工载体
结合了PAC和双元载体的优点:
1、具有多克隆位点,2、具有P1复制子和RI质粒复制子能在大肠杆菌和农杆菌中以单拷贝的形式穿梭复制。
3、其T-DNA 右边界处插入了植物选择标记潮霉素磷酸转移酶基因(hpt )和Pnos 启动子。
4、在TAC 载体中重组筛选标记是卡那霉素抗性基因和sacB 基因
应用TAC载体在获得目的克隆后可不必进行烦琐的亚克隆,而可直接进行植物转化并开展功能互补分析,可大大促进功能基因的精细定位和图位克隆。
TAC 载体具有克隆大片段DNA 和借助于农杆菌直接转化植物的功能。
TAC还具有以下优越性:
①可通过农杆菌介导直接进行基因功能互补实验。
TAC 载体中具有P1 裂解子,可以在IPTG 的诱导下产生5-20个拷贝,提高了质粒产量。
MAC-哺乳动物人工染色体
1外源DNA片段的容量大;②不整合到宿主染色体上,不会破坏宿主基因组的结合和功能,可以稳定地存在;③较少发生基因沉默现象
MAC构建的理论基础在于将哺乳动物染色体复制起始位点序列、端粒和着丝点分离出来,然后和一些载体元件组合构成人工染色体,可以将外源基因及其调控序列引入哺乳动物。
它与宿主细胞的染色体一样稳定,但并不整合人宿主基因组。
 升级会员
升级会员 